
Electronic Cigarette Exposure Increases the Severity of Influenza a Virus Infection via TRAIL Dysregulation in Human Precision-Cut Lung Slices
 ,
,
Abstract
:1. Introduction
2. Results
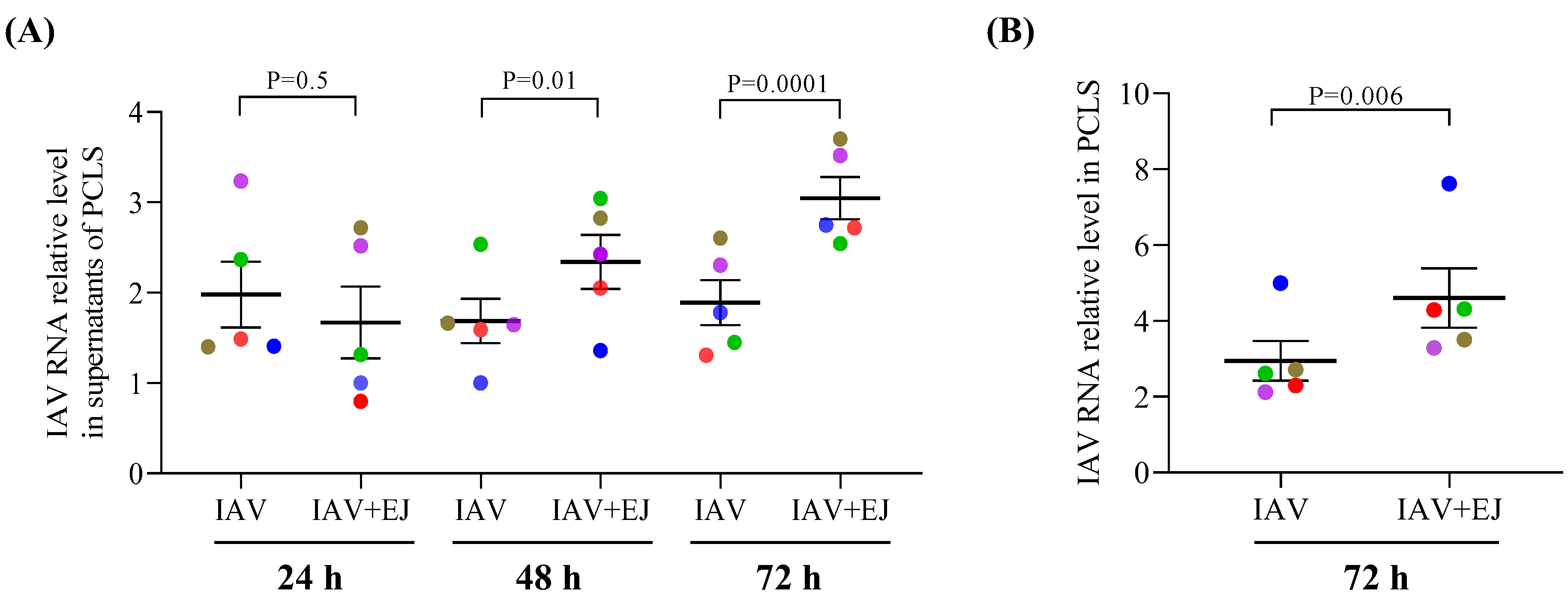
2.1. E-Juice Treatment Increased IAV Levels in Human PCLS
2.2. E-Juice Treatment Amplified TRAIL and TNF-α Release in IAV-Infected Human PCLS
2.3. E-juice Treatment Increased Cytotoxicity in IAV-Infected Human PCLS
2.4. TRAIL Neutralizing Antibody Reduced IAV Release into the Supernatant but Increased Viral Load in E-Juice Exposed Human PCLS
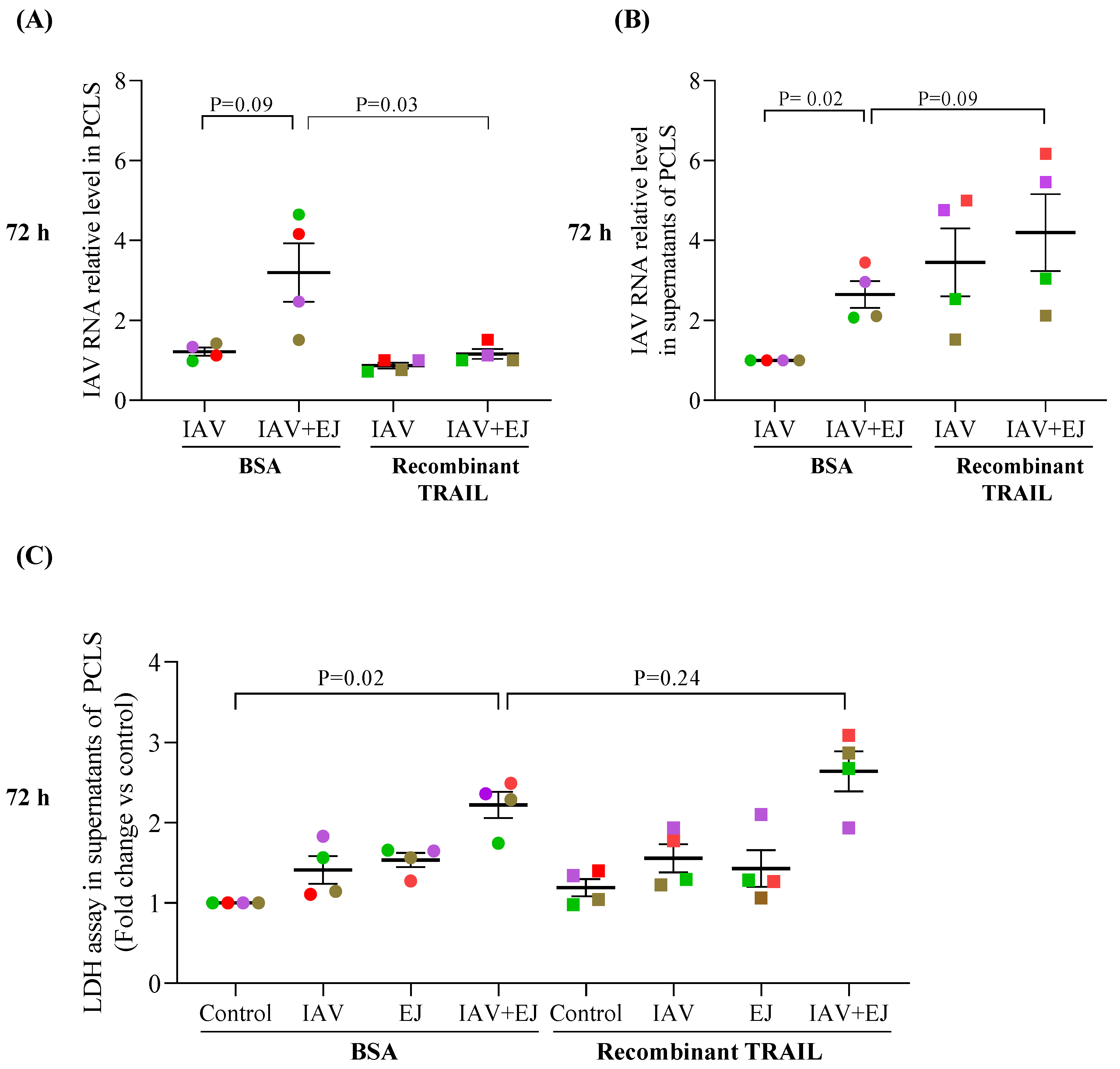
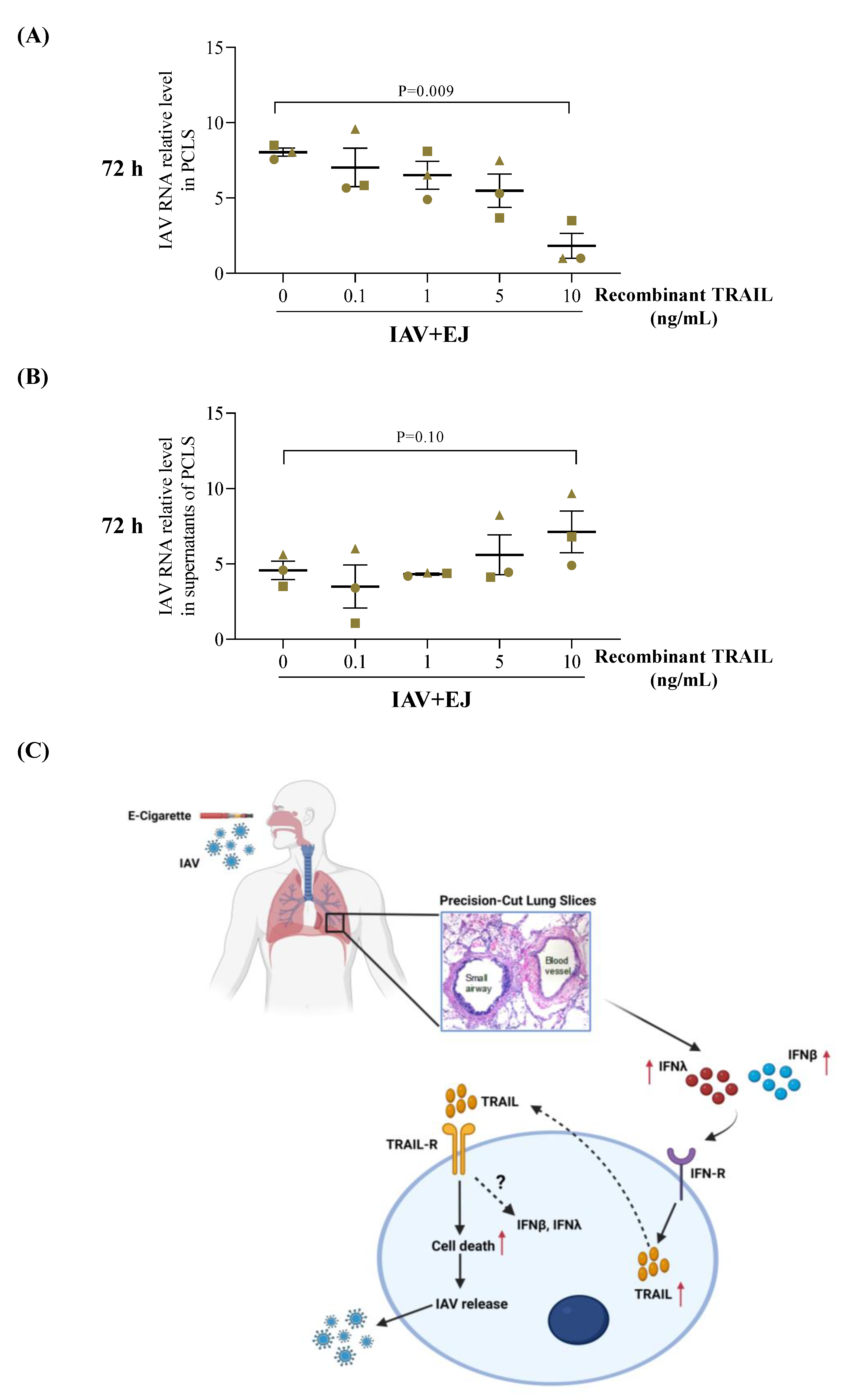
2.5. Recombinant TRAIL Decreased Tissue Viral Load but Increased IAV Release in E-Juice-Exposed Human PCLS
2.6. TRAIL Treatment Enhanced the Type I and Type III Interferon Response in PCLS Treated with E-Juice and IAV
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Preparation of PCLS from Human Donor Lungs
5.2. IAV Preparation
5.3. IAV Infection in Human PCLS
5.4. Treatments with E-Juice, TRAIL Neutralizing Antibody, and Recombinant TRAIL in Human PCLS
5.5. RT-PCR
5.6. Western Blotting
5.7. TNF-α ELISA
5.8. Lactate Dehydrogenase (LDH) Assay
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COPD | Chronic obstructive pulmonary disease |
| ALI | Air liquid interface |
| PCLS | Precision cut lung slices |
| ENDS | Electronic nicotine dispensing systems |
| E-Juice | Electronic cigarette juice |
| EJ | Electronic cigarette juice |
| EC | Electronic cigarettes |
| TRAIL | Tumor necrosis factor (TNF)-related apoptosis-inducing ligand |
| TNF-α | Tumor necrosis factor-α |
| IAV | Influenza A virus |
| EVALI | E-cigarette or vaping product use-associated lung injury |
| IFN-β | Interferon beta |
| IFN- λ | Interferon lambda |
| LDH | Lactate dehydrogenase assay |
| MDCK | Madin-Darby canine kidney |
References
- Agraval, H.; Chu, H.W. Lung Organoids in Smoking Research: Current Advances and Future Promises. Biomolecules 2022, 12, 1463. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wu, W.; Zhu, Z.; Lin, X.; Zeng, Y. The effects of epithelial–mesenchymal transitions in COPD induced by cigarette smoke: An update. Respir. Res. 2022, 23, 225. [Google Scholar] [CrossRef] [PubMed]
- Blagev, D.P.; Callahan, S.J.; Harris, D.; Collingridge, D.S.; Hopkins, R.O.; Eve, J.R.; Waddoups, L.; Aston, V.; Brown, S.; Lanspa, M.J. Prospectively Assessed Long-Term Outcomes of Patients with E-cigarette or Vaping-associated Lung Injury (EVALI). Ann. Am. Thorac. Soc. 2022, 1, 1–17. [Google Scholar] [CrossRef]
- Kalininskiy, A.; Kittel, J.; Nacca, N.E.; Misra, R.S.; Croft, D.P.; McGraw, M.D. E-cigarette exposures, respiratory tract infections, and impaired innate immunity: A narrative review. Pediatr. Med. 2021, 4, 5. [Google Scholar] [CrossRef]
- Higham, A.; Rattray, N.J.; Dewhurst, J.A.; Trivedi, D.K.; Fowler, S.J.; Goodacre, R.; Singh, D. Electronic cigarette exposure triggers neutrophil inflammatory responses. Respir. Res. 2016, 17, 56. [Google Scholar] [CrossRef] [Green Version]
- Higham, A.; Bostock, D.; Booth, G.; Dungwa, J.V.; Singh, D. The effect of electronic cigarette and tobacco smoke exposure on COPD bronchial epithelial cell inflammatory responses. Int. J. Chron. Obs. Pulmon. Dis. 2018, 13, 989–1000. [Google Scholar] [CrossRef] [Green Version]
- Putzhammer, R.; Doppler, C.; Jakschitz, T.; Heinz, K.; Förste, J.; Danzl, K.; Messner, B.; Bernhard, D. Vapours of US and EU Market Leader Electronic Cigarette Brands and Liquids Are Cytotoxic for Human Vascular Endothelial Cells. PLoS ONE 2016, 11, e0157337. [Google Scholar] [CrossRef]
- Schaunaman, N.; Crue, T.; Cervantes, D.; Schweitzer, K.; Robbins, H.; Day, B.J.; Numata, M.; Petrache, I.; Chu, H.W. Electronic cigarette vapor exposure exaggerates the pro-inflammatory response during influenza A viral infection in human distal airway epithelium. Arch. Toxicol. 2022, 96, 2319–2328. [Google Scholar] [CrossRef]
- Taha, H.R.; Al-Sawalha, N.A.; Alzoubi, K.H.; Khabour, O.F. Effect of E-Cigarette aerosol exposure on airway inflammation in a murine model of asthma. Inhal. Toxicol. 2020, 32, 503–511. [Google Scholar] [CrossRef]
- Getiye, Y.; Peterson, M.R.; Phillips, B.D.; Carrillo, D.; Bisha, B.; He, G. E-cigarette exposure with or without heating the e-liquid induces differential remodeling in the lungs and right heart of mice. J. Mol. Cell Cardiol. 2022, 168, 83–95. [Google Scholar] [CrossRef]
- Mallia, P.; Johnston, S.L. Influenza infection and COPD. Int. J. Chron. Obs. Pulmon. Dis. 2007, 2, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y.-H.; Guo, Y.; Ji, S.; Shen, J.-L.; Fei, G.-H. The Influenza A Virus H3N2 Triggers the Hypersusceptibility of Airway Inflammatory Response via Activating the lncRNA TUG1/miR-145-5p/NF-κB Pathway in COPD. Front. Pharmacol. 2021, 12, 604590. [Google Scholar] [CrossRef] [PubMed]
- Duffney, P.F.; McCarthy, C.E.; Nogales, A.; Thatcher, T.H.; Martinez-Sobrido, L.; Phipps, R.P.; Sime, P.J. Cigarette smoke dampens antiviral signaling in small airway epithelial cells by disrupting TLR3 cleavage. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2018, 314, L505–L513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viana, F.; O’Kane, C.M.; Schroeder, G.N. Precision-cut lung slices: A powerful ex vivo model to investigate respiratory infectious diseases. Mol. Microbiol. 2022, 117, 578–588. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, P.; Wang, Y.; Liu, Y.; Yang, H.; Zhou, G.; Wu, X.; Wen, Q. Application of Precision-Cut Lung Slices as an In Vitro Model for Research of Inflammatory Respiratory Diseases. Bioengineering 2022, 9, 767. [Google Scholar] [CrossRef]
- Orzalli, M.H.; Kagan, J.C. Apoptosis and Necroptosis as Host Defense Strategies to Prevent Viral Infection. Trends Cell Biol. 2017, 27, 800–809. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Lu, X.; Morken, T.; Zaki, S.R.; Katz, J.M. Depletion of lymphocytes and diminished cytokine production in mice infected with a highly virulent influenza A (H5N1) virus isolated from humans. J. Virol. 2000, 74, 6105–6116. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tan, J.; Zoueva, O.; Zhao, J.; Ye, Z.; Hewlett, I. Novel pandemic influenza A (H1N1) virus infection modulates apoptotic pathways that impact its replication in A549 cells. Microbes Infect. 2014, 16, 178–186. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Iwai, A.; Shiozaki, T.; Miyazaki, T. Relevance of signaling molecules for apoptosis induction on influenza A virus replication. Biochem. Biophys. Res. Commun. 2013, 441, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, T.; Matsukawa, S.; Higuchi, Y.; Nakamura, S.; Nakanishi, Y.; Fukuda, R. Induction of programmed cell death (apoptosis) by influenza virus infection in tissue culture cells. J. Gen. Virol. 1993, 74 Pt 11, 2347–2355. [Google Scholar] [CrossRef]
- Herold, S.; Ludwig, S.; Pleschka, S.; Wolff, T. Apoptosis signaling in influenza virus propagation, innate host defense, and lung injury. J. Leukoc. Biol. 2012, 92, 75–82. [Google Scholar] [CrossRef]
- Schneider, P.; Thome, M.; Burns, K.; Bodmer, J.L.; Hofmann, K.; Kataoka, T.; Holler, N.; Tschopp, J. TRAIL receptors 1 (DR4) and 2 (DR5) signal FADD-dependent apoptosis and activate NF-kappaB. Immunity 1997, 7, 831–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haw, T.J.; Starkey, M.R.; Nair, P.M.; Pavlidis, S.; Liu, G.; Nguyen, D.H.; Hsu, A.C.; Hanish, I.; Kim, R.Y.; Collison, A.M.; et al. A pathogenic role for tumor necrosis factor-related apoptosis-inducing ligand in chronic obstructive pulmonary disease. Mucosal Immunol. 2016, 9, 859–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Shen, Y.; Zhang, J.; Wan, C.; Wang, T.; Xu, D.; Yang, T.; Wen, F. Increased serum TRAIL and DR5 levels correlated with lung function and inflammation in stable COPD patients. Int. J. Chron. Obs. Pulmon. Dis. 2015, 10, 2405–2412. [Google Scholar] [CrossRef] [Green Version]
- Morissette, M.C.; Parent, J.; Milot, J. The emphysematous lung is abnormally sensitive to TRAIL-mediated apoptosis. Respir. Res. 2011, 12, 105. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.; Hai, R. Effects of Cigarette Smoking on Influenza Virus/Host Interplay. Pathogens 2021, 10, 1636. [Google Scholar] [CrossRef]
- Yoshimura, S.; Sano, E.; Hanashima, Y.; Yamamuro, S.; Sumi, K.; Ueda, T.; Nakayama, T.; Hara, H.; Yoshino, A.; Katayama, Y. IFN-β sensitizes TRAIL-induced apoptosis by upregulation of death receptor 5 in malignant glioma cells. Oncol. Rep. 2019, 42, 2635–2643. [Google Scholar] [CrossRef]
- Brincks, E.L.; Katewa, A.; Kucaba, T.A.; Griffith, T.S.; Legge, K.L. CD8 T cells utilize TRAIL to control influenza virus infection. J. Immunol. 2008, 181, 4918–4925. [Google Scholar] [CrossRef] [Green Version]
- Idriss, H.T.; Naismith, J.H. TNF alpha and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Churg, A.; Dai, J.; Tai, H.; Xie, C.; Wright, J.L. Tumor necrosis factor-alpha is central to acute cigarette smoke-induced inflammation and connective tissue breakdown. Am. J. Respir. Crit Care Med. 2002, 166, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Liu, T. miR-937 serves as an inflammatory inhibitor in cigarette smoke extract-induced human bronchial epithelial cells by targeting IL1B and regulating TNF-α/IL-17 signaling pathway. Tob. Induc. Dis. 2021, 19, 55. [Google Scholar] [CrossRef]
- Kumar-Sinha, C.; Varambally, S.; Sreekumar, A.; Chinnaiyan, A.M. Molecular Cross-talk between the TRAIL and Interferon Signaling Pathways*210. J. Biol. Chem. 2002, 277, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granqvist, V.; Holmgren, C.; Larsson, C. Induction of interferon-β and interferon signaling by TRAIL and Smac mimetics via caspase-8 in breast cancer cells. PLoS ONE 2021, 16, e0248175. [Google Scholar] [CrossRef] [PubMed]
- Churg, A.; Tai, H.; Coulthard, T.; Wang, R.; Wright, J.L. Cigarette Smoke Drives Small Airway Remodeling by Induction of Growth Factors in the Airway Wall. Am. J. Respir. Crit. Care Med. 2006, 174, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eapen, M.S.; Lu, W.; Hackett, T.L.; Singhera, G.K.; Mahmood, M.Q.; Hardikar, A.; Ward, C.; Walters, E.H.; Sohal, S.S. Increased myofibroblasts in the small airways, and relationship to remodelling and functional changes in smokers and COPD patients: Potential role of epithelial–mesenchymal transition. ERJ Open Res. 2021, 7, 00876–02020. [Google Scholar] [CrossRef]
- Sivaraman, V.; Parker, D.J.; Zhang, R.; Jones, M.M.; Onyenwoke, R.U. Vaping Exacerbates Coronavirus-Related Pulmonary Infection in a Murine Model. Front. Physiol. 2021, 12, 634839. [Google Scholar] [CrossRef] [PubMed]
- Bracken, M.B. Why animal studies are often poor predictors of human reactions to exposure. J. R. Soc. Med. 2009, 102, 120–122. [Google Scholar] [CrossRef] [Green Version]
- Majorova, D.; Atkins, E.; Martineau, H.; Vokral, I.; Oosterhuis, D.; Olinga, P.; Wren, B.; Cuccui, J.; Werling, D. Use of Precision-Cut Tissue Slices as a Translational Model to Study Host-Pathogen Interaction. Front. Vet. Sci. 2021, 8, 686088. [Google Scholar] [CrossRef]
- Bailey, K.E.; Pino, C.; Lennon, M.L.; Lyons, A.; Jacot, J.G.; Lammers, S.R.; Konigshoff, M.; Magin, C.M. Embedding of Precision-Cut Lung Slices in Engineered Hydrogel Biomaterials Supports Extended Ex Vivo Culture. Am. J. Respir. Cell Mol. Biol. 2020, 62, 14–22. [Google Scholar] [CrossRef]
- Alsafadi, H.N.; Uhl, F.E.; Pineda, R.H.; Bailey, K.E.; Rojas, M.; Wagner, D.E.; Königshoff, M. Applications and Approaches for Three-Dimensional Precision-Cut Lung Slices. Disease Modeling and Drug Discovery. Am. J. Respir. Cell Mol. Biol. 2020, 62, 681–691. [Google Scholar] [CrossRef]
- Rebuli, M.E.; Glista-Baker, E.; Hoffman, J.R.; Duffney, P.F.; Robinette, C.; Speen, A.M.; Pawlak, E.A.; Dhingra, R.; Noah, T.L.; Jaspers, I. Electronic-Cigarette Use Alters Nasal Mucosal Immune Response to Live-attenuated Influenza Virus. A Clinical Trial. Am. J. Respir. Cell Mol. Biol. 2021, 64, 126–137. [Google Scholar] [CrossRef]
- Sussan, T.E.; Gajghate, S.; Thimmulappa, R.K.; Ma, J.; Kim, J.H.; Sudini, K.; Consolini, N.; Cormier, S.A.; Lomnicki, S.; Hasan, F.; et al. Exposure to electronic cigarettes impairs pulmonary anti-bacterial and anti-viral defenses in a mouse model. PLoS ONE 2015, 10, e0116861. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, E.; Nakazawa, M.; Yoshinari, M.; Minami, M. Role of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand in Immune Response to Influenza Virus Infection in Mice. J. Virol. 2005, 79, 7658–7663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, N.; Badley, A. The TRAIL to viral pathogenesis: The good, the bad and the ugly. Curr. Mol. Med. 2009, 9, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.K.; Kim, J.A.; Oh, S.J.; Lee, E.W.; Shin, O.S. Zika Virus Induces Tumor Necrosis Factor-Related Apoptosis Inducing Ligand (TRAIL)-Mediated Apoptosis in Human Neural Progenitor Cells. Cells 2020, 9, 2487. [Google Scholar] [CrossRef] [PubMed]
- Romagna, G.; Allifranchini, E.; Bocchietto, E.; Todeschi, S.; Esposito, M.; Farsalinos, K.E. Cytotoxicity evaluation of electronic cigarette vapor extract on cultured mammalian fibroblasts (ClearStream-LIFE): Comparison with tobacco cigarette smoke extract. Inhal. Toxicol. 2013, 25, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Farsalinos, K.E.; Romagna, G.; Allifranchini, E.; Ripamonti, E.; Bocchietto, E.; Todeschi, S.; Tsiapras, D.; Kyrzopoulos, S.; Voudris, V. Comparison of the cytotoxic potential of cigarette smoke and electronic cigarette vapour extract on cultured myocardial cells. Int. J. Env. Res. Public Health 2013, 10, 5146–5162. [Google Scholar] [CrossRef] [Green Version]
- Cervellati, F.; Muresan, X.M.; Sticozzi, C.; Gambari, R.; Montagner, G.; Forman, H.J.; Torricelli, C.; Maioli, E.; Valacchi, G. Comparative effects between electronic and cigarette smoke in human keratinocytes and epithelial lung cells. Toxicol. Vitr. 2014, 28, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Stanifer, M.L.; Guo, C.; Doldan, P.; Boulant, S. Importance of Type I and III Interferons at Respiratory and Intestinal Barrier Surfaces. Front. Immunol. 2020, 11, 608645. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Metcalf, J.P. The Role of Type I IFNs in Influenza: Antiviral Superheroes or Immunopathogenic Villains? J. Innate Immun. 2020, 12, 437–447. [Google Scholar] [CrossRef]
- Kayagaki, N.; Yamaguchi, N.; Nakayama, M.; Eto, H.; Okumura, K.; Yagita, H. Type I Interferons (IFNs) Regulate Tumor Necrosis Factor–related Apoptosis-inducing Ligand (TRAIL) Expression on Human T Cells: A Novel Mechanism for the Antitumor Effects of Type I IFNs. J. Exp. Med. 1999, 189, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Makowska, A.; Wahab, L.; Braunschweig, T.; Kapetanakis, N.-I.; Vokuhl, C.; Denecke, B.; Shen, L.; Busson, P.; Kontny, U. Interferon beta induces apoptosis in nasopharyngeal carcinoma cells via the TRAIL-signaling pathway. Oncotarget 2018, 9, 14228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Jiang, D.; Minor, M.; Chu, H.W. Electronic cigarette liquid increases inflammation and virus infection in primary human airway epithelial cells. PLoS ONE 2014, 9, e108342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, K.M.; Moyo, N.A.; Leeming, G.H.; Bingle, L.; Jasim, S.; Hussain, S.; Schorlemmer, A.; Kipar, A.; Digard, P.; Tripp, R.A.; et al. An innate defense peptide BPIFA1/SPLUNC1 restricts influenza A virus infection. Mucosal Immunol. 2018, 11, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Madison, M.C.; Landers, C.T.; Gu, B.H.; Chang, C.Y.; Tung, H.Y.; You, R.; Hong, M.J.; Baghaei, N.; Song, L.Z.; Porter, P.; et al. Electronic cigarettes disrupt lung lipid homeostasis and innate immunity independent of nicotine. J. Clin. Investig. 2019, 129, 4290–4304. [Google Scholar] [CrossRef] [Green Version]
- Obernolte, H.; Niehof, M.; Braubach, P.; Fieguth, H.-G.; Jonigk, D.; Pfennig, O.; Tschernig, T.; Warnecke, G.; Braun, A.; Sewald, K. Cigarette smoke alters inflammatory genes and the extracellular matrix—investigations on viable sections of peripheral human lungs. Cell Tissue Res. 2022, 387, 249–260. [Google Scholar] [CrossRef]
- Herbert, J.; Kelty, J.; Laskin, J.; Laskin, D.; Gow, A. Menthol Flavoring in E-Cigarette Condensate Causes Pulmonary Dysfunction and Cytotoxicity in Precision Cut Lung Slices. Am. J. Physiol. Lung Cell Mol. Physiol. 2023. [Google Scholar] [CrossRef]
- Numata, M.; Mitchell, J.R.; Tipper, J.L.; Brand, J.D.; Trombley, J.E.; Nagashima, Y.; Kandasamy, P.; Chu, H.W.; Harrod, K.S.; Voelker, D.R. Pulmonary surfactant lipids inhibit infections with the pandemic H1N1 influenza virus in several animal models. J. Biol. Chem. 2020, 295, 1704–1715. [Google Scholar] [CrossRef]
- Chan, R.W.Y.; Yuen, K.M.; Yu, W.C.L.; Ho, C.C.C.; Nicholls, J.M.; Peiris, J.S.M.; Chan, M.C.W. Influenza H5N1 and H1N1 Virus Replication and Innate Immune Responses in Bronchial Epithelial Cells Are Influenced by the State of Differentiation. PLoS ONE 2010, 5, e8713. [Google Scholar] [CrossRef]
- Daly, K.; Nguyen, P.; Woodland, D.L.; Blackman, M.A. Immunodominance of major histocompatibility complex class I-restricted influenza virus epitopes can be influenced by the T-cell receptor repertoire. J. Virol. 1995, 69, 7416–7422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartshorn, K.L.; White, M.R.; Tecle, T.; Holmskov, U.; Crouch, E.C. Innate Defense against Influenza A Virus: Activity of Human Neutrophil Defensins and Interactions of Defensins with Surfactant Protein D. J. Immunol. 2006, 176, 6962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisaki, K.M.; Janoff, E.N. Influenza in immunosuppressed populations: A review of infection frequency, morbidity, mortality, and vaccine responses. Lancet Infect. Dis. 2009, 9, 493–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yingst, J.M.; Foulds, J.; Veldheer, S.; Hrabovsky, S.; Trushin, N.; Eissenberg, T.T.; Williams, J.; Richie, J.P.; Nichols, T.T.; Wilson, S.J.; et al. Nicotine absorption during electronic cigarette use among regular users. PLoS ONE 2019, 14, e0220300. [Google Scholar] [CrossRef]
- Marsot, A.; Simon, N. Nicotine and Cotinine Levels With Electronic Cigarette: A Review. Int. J. Toxicol. 2016, 35, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Chiadmi, F.; Schlatter, J. Simultaneous determination of cotinine and trans-3-hydroxycotinine in urine by automated solid-phase extraction using gas chromatography-mass spectrometry. Biomed. Chromatogr. 2014, 28, 453–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Gender | Age (Years) | Smoking History |
|---|---|---|---|
| 1 | Female | 32 | Non-smoker |
| 2 | Female | 27 | Non-smoker |
| 3 | Male | 30 | Non-smoker |
| 4 | Male | 73 | Non-smoker |
| 5 | Male | 63 | Non-smoker |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agraval, H.; Crue, T.; Schaunaman, N.; Numata, M.; Day, B.J.; Chu, H.W. Electronic Cigarette Exposure Increases the Severity of Influenza a Virus Infection via TRAIL Dysregulation in Human Precision-Cut Lung Slices. Int. J. Mol. Sci. 2023, 24, 4295. https://doi.org/10.3390/ijms24054295
Agraval H, Crue T, Schaunaman N, Numata M, Day BJ, Chu HW. Electronic Cigarette Exposure Increases the Severity of Influenza a Virus Infection via TRAIL Dysregulation in Human Precision-Cut Lung Slices. International Journal of Molecular Sciences. 2023; 24(5):4295. https://doi.org/10.3390/ijms24054295
Chicago/Turabian StyleAgraval, Hina, Taylor Crue, Niccolette Schaunaman, Mari Numata, Brian J. Day, and Hong Wei Chu. 2023. "Electronic Cigarette Exposure Increases the Severity of Influenza a Virus Infection via TRAIL Dysregulation in Human Precision-Cut Lung Slices" International Journal of Molecular Sciences 24, no. 5: 4295. https://doi.org/10.3390/ijms24054295